|
Von Graphen, Genen und dem WWW

Online-Skriptum
Verwandtschaft 1
Verwandtschaft
ist in der Biologie eine zentrale Kategorie. Sie stellt gewissermaßen
das Medium der Evolution dar, und ihrer Formalisierung wenden wir uns
nun zu.
Nicht nur die Evolution
von Populationen, wie sie uns in einem früheren
Abschnitt begegnet ist, auch individuelle Verwandtschaftsverhältnisse
können durch Graphen dargestellt werden. Wir nennen sie Verwandschaftsgraphen.
Formal handelt es sich um ecken- und kantengewichtete gerichtete
Graphen. Sie werden in der Praxis oft als "Stammbäume"
bezeichnet, sind aber genau genommen nicht immer "Bäume"
im graphentheoretischen Sinn. Im
Fall von Organismen, die sich nur auf sexuelle Weise fortpflanzen
können (das betrifft die meisten Tierarten), besitzen sie drei
Eckentypen und zwei Kantentypen: Zwei Eckentypen stellen
weibliche ( ) und
männliche( ) und
männliche( )
Individuen dar. Graue ausgezogene Kanten ( )
Individuen dar. Graue ausgezogene Kanten ( )
bezeichnen die Paarungen von je zwei verschiedengeschlechtlichen Individuen.
Sie enden in einem dritten Eckentyp ( )
bezeichnen die Paarungen von je zwei verschiedengeschlechtlichen Individuen.
Sie enden in einem dritten Eckentyp ( ),
der für die geschlechtliche Fortpflanzung steht. Von ihm gehen
rote ausgezogene Kanten ( ),
der für die geschlechtliche Fortpflanzung steht. Von ihm gehen
rote ausgezogene Kanten ( )
zu den von dem betreffenden Elternpaar gezeugten Kindern aus. Die Zeit
(genauer: die Generationenfolge) verläuft von oben nach unten.
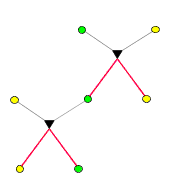
Ein typischer Verwandschaftsgraph sieht so aus: )
zu den von dem betreffenden Elternpaar gezeugten Kindern aus. Die Zeit
(genauer: die Generationenfolge) verläuft von oben nach unten.
Ein typischer Verwandschaftsgraph sieht so aus:
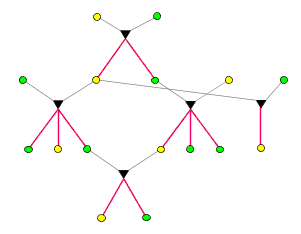
Hier sind von
insgesamt 11
Individuen die unmittelbar vorausgehenden Abstammungsverhältnisse
dargestellt. Auch die
5 Elternteile,
die gewissermaßen vom Himmel fallen, stammen natürlich ihrerseits
von Eltern ab, werden aber als nicht miteinander verwandt betrachtet:
Als verwandt gilt nur, wer das auf der Basis des Graphen ist. Verwandschaftsgraphen
erlauben die Darstellung ganz allgemeiner Familienverhältnisse.
So gibt es hier beispielsweise Kinder, die denselben Vater, aber verschiedene
Mütter haben (Halbgeschwister). Weiters ist eine Verbindung von
Cousin und Cousine ("Vetternehe"), die zwei Kinder hervorgebracht hat,
dargestellt.
An jeder -Ecke
findet für jedes Kind die Rekombination der Erbanlagen gemäß
den im vorigen Abschnitt besprochenen Mendelschen
Gesetzen statt. Alle Kanten entsprechen dem Vorhandensein diploider
Erbinformation, d.h. das Erbmaterial liegt in Form von Allelpaaren
vor. (Bienen, Wespen und Ameisen machen es ein bisschen anders -
wir werden weiter unten
darauf zu sprechen kommen). Beschränken wir uns auf einen Genort,
so läuft an jeder -Ecke
ein einfacher Zufallsprozess ab: Jedes Kind bekommt ein Allel von Vater
und eines von der Mutter - welches, das bestimmt
der Zufall. Jedes Allel, das ein Elternteil trägt, wird mit einer
Wahrscheinlichkeit von 1/2
auf das Kind vererbt. Dabei wird die Möglichkeit, dass es zu eineiigen
Zwillingen (genetisch identischen Geschwistern) kommt, vernachlässigt.
Die Einfachheit dieser Vorgänge erlaubt es, Verwandtschaftsstrukturen
zu formalisieren. Dazu benötigen wir zunächst ein paar Definitionen:
- Wie bereits erwähnt,
handelt es sich bei den Verwandtschaftsgraphen um gerichtete Graphen,
obwohl wir uns das Zeichnen von Pfeilen ersparen: Jede Kante (beiderlei
Typs) weist von den Vorfahren zu den Nachkommen, was wir als absteigend
bezeichnen. Wollen wir eine Kante in umgekehrter Richtung durchlaufen,
so nennen wir sie aufsteigend.
- Sei x
ein Individuum (d.h. eine Ecke des Typs
oder ). Ein Nachkomme
von x
ist ein (von x
verschiedenes) Individuum, das von x
über einen absteigenden Kantenzug erreicht werden kann. Ein Vorfahr
von x
ist ein (von x
verschiedenes) Individuum, das von x
über einen aufsteigenden Kantenzug erreicht werden kann.
- Zwei Individuen x
und y
sind miteinander verwandt, wenn sie (innerhalb des Graphen)
zumindest einen gemeinsamen Vorfahren besitzen.
- Ein Individuum heißt
Gründer, wenn von ihm keine aufsteigende Kante ausgeht,
d.h. wenn seine Eltern nicht im Diagramm erfasst sind. (Wir legen
fest, dass für alle anderen Individuen beide Eltern im
Graphen enthalten sein sollen). Alle Gründer gelten als untereinander
nicht verwandt.
- Von Inzucht (Inzest,
inbreeding) sprechen wir ganz allgemein, wenn Verwandte Nachkommen
zeugen. In einem Verwandtschaftsgraphen erweist sie sich durch die
Existenz zumindest eines nicht-trivialen Kreises, der aus einem
aufsteigenden und einem absteigenden Kantenzug besteht.
- Liegt in einem Graphen keine
Inzucht vor, so nennen wir ihn inzuchtfrei. Enthält ein
Graph keinen Kreis, so nennen wir ihn baumartig. (Wir verlangen
nicht, dass er zusammenhängend ist -
er stellt also nicht notwendigerweise einen "Baum" im graphentheoretischen
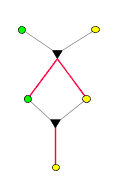
Sinn dar). Jeder baumartige Graph ist inzuchtfrei. Hier drei paar
Beispiele, deren mittleres zeigt, dass nicht jeder inzuchtfreie Graph
baumartig ist:
 |
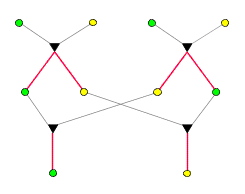 |
 |
| baumartig |
nicht
baumartig,
inzuchtfrei |
nicht
baumartig,
nicht inzuchtfrei |
Ein Graph ist genau dann inzuchtfrei, wenn jedes Individuum mit jedem
Gründer durch höchstens einen aufsteigenden Kantenzug
verbunden werden kann.
(Beweis: Gäbe es zwei solcher Kantenzüge von x
zu einem Gründer - die sich o.B.d.A.
unmittelbar oberhalb von x
trennen -, so wäre dieser Gründer
ein gemeinsamer Vorfahr der Eltern von x,
letztere also miteinander verwandt. Sind umgekehrt die Eltern von
x
miteinander verwandt, so kann man von einem ihrer gemeinsamen Vorfahren
bis zu einem Gründer aufsteigen und hätte damit zwei
verschiedene Wege von x
zu diesem Gründer konstruiert).
- Ein Verwandtschaftsgraph
ist genau dann baumartig, wenn je zwei verwandte Individuen durch
genau einen Weg W
(d.h. einen Kantenzug, dessen Kanten alle voneinander verschieden
sind) verbunden werden können (Beweis: Übungsaufgabe). W
besitzt eine eindeutig bestimmte Umkehr-Ecke, die (falls sie vom Typ
oder
ist) den jüngsten gemeinsamen Vorfahren oder (falls sie vom Typ
ist) das jüngste gemeinsame
Vorfahrenpaar darstellt. Dank dieser Eigenschaft sind baumartige Verwandtschaftsgraphen
besonders leicht zu behandeln.
Für zahlreiche
Zwecke (vom Zuchtmanagement zum Schutz bedrohter Arten über die
medizinische Forschung bis zu Grundsatzfragen der Evolutionstheorie)
benötigt die Populationsgenetik ein quantitatives Maß für
die Nähe der Verwandtschaft zweier Individuen und für die
Gefahren, die Inzucht mit sich bringt.
Um den Grad der
Verwandschaft zu formalisieren, beschränken wir uns auf einen
Genort und nehmen an, dass genau ein Gründer an diesem Genort
ein Allel (in einfacher Ausführung) trägt, das von
allen Allelen der anderen Gründer verschieden ist. (Wir nennen
es ein "seltenes Allel". Da es nur in einfacher Ausführung
vorliegt, ist der betreffende Gründer an diesem Genort heterozygot).
Für jeden Gründer besteht die gleiche Wahrscheinlichkeit,
es zu besitzen. Der Sinn dieses formalen Tricks besteht darin, das seltene
Allel gewissermaßen als "Markierung" im Fluss der Gene
zu benutzen und seinen Werdegang im Graphen zu verfolgen. (Wir können
uns auch vorstellen, dass es in einem Gründer durch Mutation entstanden
ist). Tritt es in irgendeinem Individuum auf, ist klar, dass es letztlich
von genau einem Gründer stammt. Tritt es in zwei verschiedenen
Individuen auf, so tut es das aus Gründen der gemeinsamen Abstammung.
Da der Formalismus
der Verwandtschaft in baumartigen (und teilweise auch in inzuchtfreien)
Verwandtschaftsgraphen recht einfach ist, besprechen wir diesen zuerst.
Im nächsten Abschnitt werden
wir uns allgemeinen Verwandtschaftsgraphen zuwenden.
Betrachten wir
einen beliebigen baumartigen Verwandtschaftsgraphen. Wer ist mit wem
"wie nahe" verwandt? Wichtig ist zunächst, dass in jedem
inzuchtfreien (also auch in jedem baumartigen) Verwandtschaftsgraphen
jedes Individuum das seltene Gen höchstens einfach trägt.
(Es kann nur an die Nachkommen des Gründers, der es eingebracht
hat, vererbt werden. Da stets nur nichtverwandte Partner gewählt
werden, kann das seltene Allel jedem Individuum nur entlang einer einzigen
Linie übertragen werden, und zwar immer nur in einfacher Ausführung).
Jedes Induviduum trägt es oder trägt es nicht. Das macht die
Analyse inzuchfreier Verwandtschaftsgraphen so einfach.
Seien nun x
und y
zwei Individuen (d.h. Ecken in einem Verwandtschaftsgraphen). Falls
x
das seltene Allel besitzt, bezeichnen wir mit P(y|x)
die Wahrscheinlichkeit, dass es y
auch besitzt. (Es ist dies eine bedingte Wahrscheinlichkeit: der senkrechte
Strich |
wird am besten als "vorausgesetzt, dass" gelesen). P(y|x)
wäre auch schon ein Kandidat für ein quantitatives Maß
der Nähe der Verwandtschaft. Wir können aber auch die Rollen
von x
und y
vertauschen und statt dessen P(x|y)
betrachten. Interessanterweise sind diese beiden Größen in
inzuchtfreien Verwandtschaftsgraphen immer gleich. (In beliebigen
Verwandtschaftsgraphen gilt das nicht!) Unter Benutzung der Schreibweisen
- P(x) =
Wahrscheinlichkeit, dass x
das seltene Allel besitzt
- P(y) =
Wahrscheinlichkeit, dass y
das seltene Allel besitzt
- P(x, y) =
Wahrscheinlichkeit, dass x
und y
das seltene Allel besitzen
fassen wir einige
wichtige Tatsachen zusammen:
Satz 1:
In einem inzuchtfreien Verwandtschaftsgraphen gilt
- Ist g
die Anzahl der Gründer, so ist P(x) = 1/g
für alle Individuuen x.
- P(x|y) = P(y|x) = g P(x, y)
für je zwei Individuen x
und y.
- Ist der Graph baumartig,
so betrachten wir den (eindeutig bestimmten) Kantenzug W,
der aufsteigend in x
beginnt, beim jüngsten gemeinsamen Vorfahren(paar) umkehrt, absteigend
in y
endet und dabei keine Kante zweimal durchläuft. Sei k
der "Generationenabstand" zwischen x
und y,
d.h. die Anzahl der Ecken vom Typ ,
die W
enthält. Dann ist P(x|y) = P(y|x) = 2-k.
Beweis:
- Diese Eigenschaft überträgt sich von den Eltern auf
jedes Kind. Für die Gründer gilt sie per Definition. Sind
nun u
und w
die Eltern von x,
so nehmen wir an, dass die Aussage für die Eltern gilt und
zeigen, dass sie auch für das Kind x
zutrifft: Da es sich um einen inzuchtfreien Graphen handelt,
können u
und w
nicht beide das seltene Allel tragen (ansonsten wären
sie miteinander verwandt). Gemäß dem Mechanismus der
sexuellen Fortpflanzung ist die Wahrscheinlichkeit, dass x
das Allel von u
erbt, 1/(2g),
und die Wahrscheinlichkeit, dass es das Allel von
w erbt, ebenfalls 1/(2g).
Insgesamt wird x
das seltene Allel mit der Wahrscheinlichkeit 1/g
erben. Damit ist Punkt 1 bewiesen.
- Es handelt sich um eine triviale Folgerung aus 1 und der Definition
der bedingten Wahrscheinlichkeit:
P(x, y) = P(x) P(y|x) = P(y) P(x|y).
- Wir nehmen an, dass
x das seltene Allel besitzt. P(y|x)
ist die Wahrscheinlichkeit, dass y
es auch besitzt. Wir wandern den (aufgrund der Baumartigkeit eindeutig
bestimmten) Weg W
von x
nach y
und benutzen die aus Punkt 1 folgende Tatsache, dass jeder Elternteil
eines Individuums das Allel mit der gleichen Wahrscheinlichkeit
trägt. Die Wahrscheinlichkeit, dass x
das Allel von jenem Elternteil u
geerbt hat, der auf W
liegt, ist 1/2.
(Das folgt unmittelbar aus der unter Punkt 2 genannten Symmetrie:
P(u|x) =
P(x|u) = 1/2). Die Wahrscheinlichkeit,
dass u
das Allel von jenem Elternteil v
geerbt hat, der auf W
liegt, ist ebenfalls 1/2,
und so weiter. Das machen wir, bis wir bei der Umkehr-Ecke w
angelangt sind. Danach steigen wir zu y
ab: Die Wahrscheinlichkeit, dass jenes der Kinder von w,
das auf W
liegt, das Allel erbt, ist 1/2,
und so weiter. Insgesamt ist die Wahrscheinlichkeit, dass y
das Allel erbt, 2-k,
wobei k
die Anzahl der "Stationen" (der -Ecken)
auf dem Weg von x
nach y
war.
Definition:
Als Verwandtschaftsgrad (coefficient of relationship)
zwischen zwei Individuen x
und y
in einem inzuchtfreien Verwandtschaftsgraphen wird die Größe
| r(x, y) = P(x|y) = P(y|x) |
(1) |
bezeichnet. Da
P(x, y) = P(x) P(y|x) = P(y) P(x|y)
gilt, kann der Verwandschaftsgrad auch in der Form
| r(x, y) = |
P(x, y)
|
= |
P(x, y)
|
|
| P(x) |
P(y) |
|
(2) |
geschrieben werden.
Er hat also eine intuitiv einleuchtende Bedeutung: Besitzt x
das seltene Allel, so ist r(x, y)
die Wahrscheinlichkeit dafür,
dass y
es auch besitzt. Aus Satz 1
folgt, dass in dieser Aussage x
und y
ihre Rollen vertauschen können. r(x, y)
liegt immer zwischen 0
und 1. Die
beiden Extremfälle sind:
- Den höchsten
Verwandtschaftsgrad 1
haben nur Individuen, die (aufgrund des Graphen) genetisch identisch
sind. Das trifft (außer beim Vergleich eines Individuums mit
sich selbst) nur auf eineiige Zwillinge zu. Vom genetischen Standpunkt
müssen sie nicht unterschieden werden. In den folgenden Berechnungen
gehen wir davon aus, dass die in einem Graphen dargestellten Geschwister
keine eineiigen Zwillinge sind. Formal gilt für jedes
Individuum r(x, x) = 1.
- Der niedrigste Verwandtschaftsgrad
0
besteht zwischen zwei Individuen genau dann, wenn sie (aufgrund des
Graphen) als nicht miteinander verwandt gelten.
Für Verwandtschaftsgrade
in baumartigen Graphen haben wir mit Punkt 3 von Satz 1
eine bequeme Berechnungsmethode. (Methoden, die in beliebigen
Graphen anwendbar sind, werden wir im nächsten
Abschnitt besprechen). Sehen wir uns einige Beispiele an:
- Verwandtschaftsgrad zwischen
Geschwistern:
Die Anwendung von Satz 1,
Punkt 3 ergibt sofort, dass der Verwandtschaftsgrad zwischen Geschwistern
1/2 ist
(sofern die Eltern als nicht miteinander verwandt gelten).
- Verwandtschaftsgrad zwischen
Eltern und Kindern:
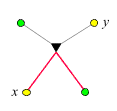
Der Verwandtschaftsgrad ist ebenfalls 1/2.
Eltern sind mit ihren Kindern genauso nahe verwandt wie Geschwister
untereinander.
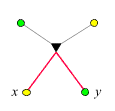
- Verwandtschaftsgrad zwischen
Tante/Onkel und Nichte/Neffe:
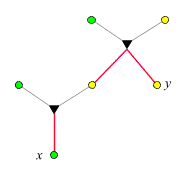
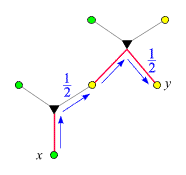
Der Verwandtschaftsgrad ist 1/4,
da auf dem Weg von x
nach y zwei
-Ecken
liegen. Wir nutzen die Gelegenheit und exerzieren zum besseren Verständnis
die Logik des obigen
Beweises (Punkt 3) anhand dieses Beispiels vor: x
besitze das seltene Allel. Wir wandern in der im rechten Bild dargestellten
Weise von x
nach y
und schreiben jedesmal, wenn sich eine Aufspaltung ergibt, die entsprechende
Wahrscheinlichkeit dazu: Da x
das seltene Allel besitzt, und da es nur entlang eines Kantenzugs
auf x
übergegangen sein kann, wurde es mit Wahrscheinlichkeit
1/2 vom
Vater übertragen. (Das folgt unmittelbar aus P(v|x) =
P(x|v) = 1/2, wobei v
den Vater von x
bezeichnet). War das der Fall, so trug es genau ein Elternteil von
y.
Die Wahrscheinlichkeit, dass es dann auch auf y
übertragen wurde, ist 1/2.
Insgesamt ist die Wahrscheinlichkeit, dass y
es erbt, 1/4.
Jede -Ecke
auf dem Weg von x
nach y
trägt einen Faktor 1/2
bei.
- Verwandtschaftsgrad zwischen
Cousins/Cousinen (ersten Grades):
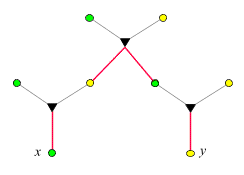
Nach derselben Regel ergibt sich ein Verwandtschaftsgrad von 1/8.
- Der Verwandtschaftsgrad zwischen
Halbgeschwistern
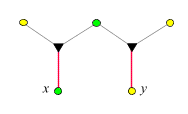
ergibt sich nach derselben Regel zu 1/4.
Es sei nochmals betont, dass
alle diese Resultate darauf beruhen, dass die Gründer als nicht
miteinander verwandt angesehen werden, d.h. dass frühere Verwandtschaftsverhältnisse,
die außerhalb des Graphen liegen, nicht berücksichtigt werden.
Was können wir mit dem
Begriff des Verwandtschaftsgrades anfangen? Sehen wir uns ein Beispiel
an, das unser Grundverständnis der Evolution betrifft.
| Verwandtschaftsaltruismus
und das "egoistische Gen" |
Eine vieldiskutierte
Fragen der modernen Evolutionsbiologie lautet: Wie ist Altruismus
entstanden, d.h. die Bereitschaft, andere Individuen auf Kosten des
eigenen Fortpflanzungserfolgs zu unterstützen? Der Hinweis, dass
Altruismus "für die Art" nützlich sei, hilft nicht
wirklich weiter, da er nicht erklärt, wie sich altruistische Anlagen
durchsetzen (d.h. in einer Population ausbreiten) können, solange
sie noch in der Minderheit sind. Interessanterweise hat hier die Mathematik
ein Wort mitzureden.
Um die Struktur
des Problems zu verstehen, betrachten wir ein stark vereinfachtes Szenario:
Wir nehmen an, es entstünde durch Mutation in einer Population
ein Gen, das seine Träger dazu veranlasst, unter gewissen Umständen
mit anderen Individuen zu kooperieren. Das neue Gen gehört zu einem
bestimmten Genort, tritt also als Allel auf, das mit anderen Allelen
(die nicht zu Altruismus führen) um die Besetzung dieses Genorts
konkurriert, und zunächst wird es nur von wenigen Individuen getragen.
Wird es sich in der Population ausbreiten? Dazu ist zunächst festzuhalten:
Allele haben natürlich keine Motive, aber in einem übertragenen
Sinn lässt sich sagen, dass es dem Allel gleichgültig ist,
welche Individuen sich fortpflanzen. Es bezieht seinen Erfolg
lediglich daraus, dass die Zahl seiner im Umlauf befindlichen Kopien
von Generation zu Generation wächst. Das ist mit dem populären
Schlagwort vom "egoistischen Gen" gemeint.
Hier sind
einige Bemerkungen angebracht. In der Evolutionsbiologie geht es weder
darum, für alles "die Gene" verantwortlich zu machen,
noch um das Postulieren hypothetischer Genen für jede Verhaltensform.
Tatsächlich ist konkretes Verhalten immer vom Milieu mibestimmt,
und die einer Art eigenen Verhaltensmuster, soweit sie genetischen
Ursprungs sind, werden von vielen Genen (so genannten "Genkomplexen")
geregelt, die in komplexer Weise zusammenspielen. Allerdings ist auch
das Geschehen auf der Ebene einzelner Gene wichtig:
- Liegen
mehrere Genorte auf einem Chromosom nahe beieinander, so werden
die auf ihnen sitzenden Allele mit großer Wahrscheinlichkeit
bei der Rekombination nicht getrennt, treten daher über viele
Generationen in derselben Kombination auf. Für solche Allelkombinationen
gelten näherungsweise dieselben Vererbungsregeln wie für
einzelne Allele. Darin besteht für die Evolution eine Chance,
Information eines gewissen Komplixitätsgrads vor ständiger
Durchmischung zu bewahren.
- Einzelne
(oder wenige) Allele können maßgeblich dafür sein,
ob die an bestimmten anderen Genorten befindlichen Instruktionen
überhaupt ausgeführt werden. Diese "Schalter"
können komplexe Informationen, die im Genom einer Art vorhanden
sind, mit einem Schlag modifizieren, z.B. an Bedingungen knüpfen.
Auf diese Weise kann die Evolution Altes umformen, für neue
Zwecke einsetzen und Ungünstiges auf Eis legen.
- Auch wenn
eine neue Allelvariante nur einen geringfügigen Vorteil gegenüber
ihren alteingesessenen Konkurrenten hat, kann sie im Laufe der Zeit
von der natürlichen Selektion stark bevorzugt werden.
Das
sollte nicht vergessen werden, wenn im Folgenden vereinfachend von einem
"Altruismus-Allel"
gesprochen wird.
Wenn nun etwa Altruist
auftritt, der anderen Individuen wahllos bei der Behütung des Nachwuchses
hilft, stellt sich die Frage, ob das Allel, das ihn dazu veranlasst,
etwas davon hat. Das wird im Allgemeinen nicht der Fall sein. Bessere
Aussichten hätte eine Allel, das Kooperation mit anderen Trägern
desselben Allels nahelegt! Das
Problem bei einer solchen Variante wäre, wie ein anderer Träger
des Altruismus-Allels erkannt werden kann. Eine mögliche Antwort
ist: durch Verwandtschaft, die sich -
auch für Tiere - durch die Umständen,
unter denen man aufwächst, erschließt. Nehmen
wir beispielsweise an, unser Allel steht für folgende Instruktion
an seinen Träger: "Wenn Nahrungsmangel herrscht und deine
Schwester bereits Kinder hat, so verzichte auf eigene Nachkommen und
sei ihr behilflich!" Individuum x
sei Träger des Allels und finde die genannten Bedingungen vor.
Ihre Schwester hat zwei Kinder. Wie groß ist die Wahrscheinlichkeit
für ein Kind, dass es das Altruismus-Allel ebenfalls trägt?
Das ist genau der Verwandtschaftsgrad, den wir oben
definiert haben! Zwischen Tante/Onkel und Nichte/Neffe beträgt
er, sofern früherere Verwandtschaftsverhältnisse ignoriert
werden, 1/4
(ansonsten ist er der Regel noch größer). Die Wahrscheinlichkeit,
dass zumindest eines der Kinder das Allel trägt, ist immerhin 7/16.
Wenn die Chance, die Kinder der Schwester mit vereinten Kräften
durchzubringen, größer ist als die Aussicht, dass von zwei
Familien mit je einer Ernährerin zumindest eine die mageren Zeiten
überlebt, wird der Anteil an Trägern des Altruismus-Allels
in der Population zunehmen. Das ist die Grundidee. Evolutionsbiologen
kennen eine Reihe von Beispielen, die nach diesem Muster zu funktionieren
scheinen. So ist es etwa keineswegs bei allen Tieren üblich, allen
bedrohten Artgenossen in Gefahr beizustehen und sein Leben für
sie einzusetzen. Bei vielen Arten gilt ein solcher Einsatz vorwiegend
den eigenen Verwandten, und er scheint auf die Nähe der Verwandtschaft
recht fein abgestimmt zu sein.
Und diese Idee
formalisieren zu können, betrachten wir folgendes (natürlich
stark vereinfachtes) Modell: Für die Bereitschaft, eine gewisse
Anzahl von Geschwistern zu retten, sei ein Allel verantwortlich. Können
wir voraussagen, ob es profitieren oder untergehen würde? Wenn
der Altruist bei einer Rettungsaktion den Tod findet, geht eine Kopie
des Altruismus-Allels verloren. Wieviele Kopien des Allels werden aber
zum Ausgleich (im Durchschnitt) dabei gerettet? Das hängt von der
Anzahl der Geretteten und von der Wahrscheinlichkeit ab, dass ein Geretteter
das Allel ebenfalls trägt (d.h. vom Verwandtschaftsgrad zu den
Geretteten = 1/2
im Fall von Geschwistern). Beispielsweise:
| Rettung
von |
durchschnittliche
Anzahl
geretteter Kopien des Allels |
Erfolg
des Allels
in der Population |
| 1 Geschwister |
1/2 |
schlecht |
| 2 Geschwister |
1 |
neutral |
| 3 Geschwister |
3/2 |
gut |
Demnach hätte
ein Allel, das den Träger dazu veranlasst, für die Rettung
mindestens dreier Geschwister den eigenen Tod in Kauf zu nehmen,
in der Population Erfolg (vorausgesetzt natürlich, diese Gefahrensituationen
treten oft auf). Oder, etwas weniger bürokratisch ausgedrückt:
der Einsatz für genügend viele genügend nahe Vewandte
lohnt sich - nicht für den Getöteten,
sondern für das Altruismus-Allel. Natürlich müsste auch
der voraussichtliche zukünftige Fortpflanzungserfolg der einzelnen
Individuen (also etwa deren Alter) mit berücksichtigt werden, aber
das würde nichts Grundsätzliches an diesem Mechanismus ändern.
Tatsächlich stehen viele tierische Verhaltensweisen im Verdacht,
genau dieser Logik zu gehorchen, d.h. das Ausmaß an Kooperation
- mit fast mathematischer Präzision
- vom Verwandtschaftsgrad abhängig zu
machen.
Wir wollen noch
erwähnen, dass sich altruistische Hilfe an Nicht-Verwandte auch
auszahlen kann. So könnte etwa ein neues Allel seine Träger
dazu veranlassen, jenen Individuen Körperpflege (zur Befreiung
von Parasiten) angedeihen zu lassen, von denen man diese Hilfe selbst
einmal bekommen hat, Verweigerung aber mit Verweigerung zu vergelten
(tit for tat). Wie Computersimulationen zeigen, hat ein Allel,
das zu einer solchen Art von Verhalten führt, ebenfalls gute Erfolgschancen.
Nachdem sie einmal eine gewisse Verbreitung gefunden hat, führt
diese Verhaltensweise de facto zu einem Erkennen von Trägern desselben
Allels, auch wenn man mit ihnen nicht (auf offensichtliche Weise) verwandt
ist. Tatsächlich wurden bei Tieren auch Verhaltensweisen, die diesem
Muster zu entsprechen scheinen, beobachtet.
| Bienen,
Wespen und Ameisen |
Zu den Tierarten,
in denen es die aufopferndsten Formen der Kooperation gibt, gehören
die Bienen, Wespen und Ameisen, die zur Ordnung der Hautflügler
(Hymenoptera) zählen. Kasten steriler Weibchen (Arbeiterinnen)
verzichten auf eigenen Nachwuchs, kümmern sich um die Brut und
nehmen mannigfaltige Aufgaben wahr. Kamikaze-Bienen begehen beim Stechen
der Honigdiebe praktisch sicheren Selbstmord. Die Staaten solcher Tiere
funktionieren so perfekt wie Organismen -
inklusive eines ausgeklügelten Nachrichtensystems, der Erkennung
und Eliminierung von Eindringlingen, Temperaturregelung usw. Wie konnte
eine derart extreme Form des Altruismus entstehen? Eine mögliche
Antwort lautet, dass es sich dabei um eine spezielle Form des Verwandtschaftsaltruismus
handelt.
Bei Hautflüglern
ist die Fortplflanzung und die Rekombination von Erbmaterial anders
organisiert als bei den meisten anderen Tieren. Die tatsächlichen
Verhältnisse sich reichlich kompliziert -
wir geben hier nur die Grundzüge wieder: Zwar sind die Hautflügler-Weibchen,
wie es sich gehört, diploid (d.h. sie besitzen an jedem
Genort zwei Allele), die Männchen aber sind haploid (d.h.
sie besitzen an jedem Genort nur ein Allel). Die meisten Weibchen
werden (aufgrund der ihnen angediehenen Nahrung) unfruchtbar. Im Normalfall
sorgt lediglich die Königin für Nachwuchs. Sie wurde von einem
oder wenigen Männchen (Drohnen) befruchtet -
wir wollen hier nur den Fall betrachten, dass es sich um ein einziges
Männchen handelt - und kommt mit den
empfangenen Samenzellen ihr Leben lang aus.
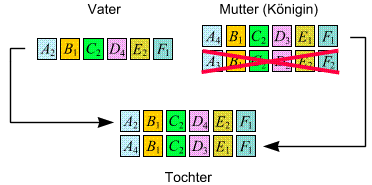
Töchter und
Söhne kommen auf auf folgende Weise zustande:
- Töchter: Das
Erbmaterial der Königin und des Männchens, das sie befruchtet
hat, rekombinieren. Die Tochter bekommt den (einzigen) Chromosomensatz
(Informationsstrang) des Vaters und einen (zufällig ausgewählten)
Satz der Königin (wobei die beiden Stränge der Königin
zuvor durch crossing over noch ein bisschen durcheinandergeschüttelt
werden):
- Söhne: Sie entstehen
in wohldosierten Mengen aus unbefruchteten Eiern und bekommen daher
lediglich einen (zufällig ausgewählten) Chromosomensatz
der Königin:
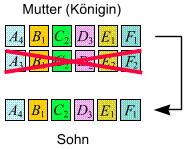
Vater ist dabei keiner vonnöten! (Da die Königin aber einen
Vater hat, besitzt jedes Männchen einen Großvater mütterlicherseits,
klarerweise aber keinen Großvater väterlicherseits!)
Für diese
Arten gelten die Mendelschen Gesetze
in abgewandelter Form: So vererbt ein Männchen jedes seiner
Allele mit Sicherheit seiner Tochter. Die Wahrscheinlichkeit, dass sich
ein von der Königin getragenes seltenes Allel in einem ihrer Kinder
wiederfindet, ist 1/2. Um diese Formen der Vererbung in einem Verwandtschaftsgraphen
darstellen zu können, führen wir zwei neue Graphenelemente
ein: Eine strichlierte Kante ( )
steht für haploide Erbinformationen, also (bei Beschränkung
auf einen Genort) für ein einziges Allel. Eine Ecke vom Typ )
steht für haploide Erbinformationen, also (bei Beschränkung
auf einen Genort) für ein einziges Allel. Eine Ecke vom Typ  repräsentiert die zufällige Auswahl eines haploiden (einfachen)
Informationssatzes aus einem diploiden (doppelten). So also kommen die
Kinder der Hautflügler zustande:
repräsentiert die zufällige Auswahl eines haploiden (einfachen)
Informationssatzes aus einem diploiden (doppelten). So also kommen die
Kinder der Hautflügler zustande:
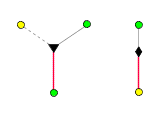
Damit sind auch
die formalen Regeln, nach denen Verwandtschaftsgraphen für diese
Arten von Insekten aufgebaut werden können, festgelegt.
Wie sieht es nun
um die Nähe der Verwandtschaft zwischen Eltern und Geschwistern
aus? Dazu benutzen wir wieder den Trick mit dem seltenen Allel. Nur
müssen wir hier ein bisschen aufpassen: Ist
in einem Verwandtschaftsgraphen
gw die Anzahl der weiblichen Gründer
und die
gm die Anzahl der männlichen Gründer,
so gibt es unter den Gründern
n = 2gw + gm
Allele, von denen eines (mit Wahrscheinlichkeit 1/n)
als "seltenes Allel" auftritt. Bezeichnen
wir wieder mit P(x)
die Wahrscheinlichkeit für ein Individuum x,
das seltene Allel zu tragen, so gilt
Satz 2:
In einem inzuchtfreien Hautflügler-Graphen gilt für
jedes Individuum x
| P(x) = |
{ |
2/n,
wenn x
weiblich ist, |
| 1/n,
wenn x
männlich ist. |
|
(3) |
Beweis:
Übungsaufgabe!
Dieser Satz ist
eine Modifikation von Punkt 1 von Satz 1
für die vorligene Art von Graphen. Eine unmittelbare Konsequenz
daraus ist, dass die bedingten Wahrscheinlichkeiten P(x|y)
und P(y|x)
nur dann übereinstimmen, wenn x
und y
das gleiche Geschlecht haben. (Ansonsten unterscheiden sie sich durch
einen Faktor 2).
Wir können daher den Verwandtschaftsgrad r(x, y)
zweier Individuen x
und y
gleichen Geschlechts als P(y|x)
definieren, d.h. als die Wahrscheinlichkeit, dass y
ein seltenes Allel besitzt, vorausgesetzt, dass
x es trägt. Im Fall von Individuen verschiedenen Geschlechts
gibt P(y|x)
die Nähe der Verwandtschaft "aus der Sicht von x"
und P(x|y)
die Nähe der Verwandtschaft "aus der Sicht von y"
an.
Im Allgemeinen
sind die Verhältnisse einer Generation des Staates durch diesen
Graphen
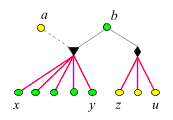
charakterisiert.
Die Berechnung der bedingten Wahrscheinlichkeiten ist nun eine einfache
Übung in Wahrscheinlichkeitsrechnung (Übungsaufgabe). Es ergibt
sich
- r(x, y) = 3/4,
d.h. Schwestern sind untereinander näher verwandt als beim Menschen.
- r(z, u) = 1/2,
d.h. Brüder sind untereinander ebenso nahe verwandt wie beim
Menschen.
- P(z|x) = 1/4,
d.h. aus der Sicht der Schwestern sind die Brüder eher entfernte
Verwandte.
- P(x|z) = 1/2,
d.h. aus der Sicht der Brüder sind die Schwestern ebenso nahe
verwandt wie beim Menschen.
- r(x, b) = 1/2,
d.h. Mutter und Töchter sind zueinander ebenso nahe verwandt
wie beim Menschen.
- P(z|b) = 1/2,
d.h. aus der Sicht der Mutter sind die Söhne ebenso nahe verwandt
wie beim Menschen.
- P(b|z) = 1,
d.h. aus der Sicht der Söhne ist die Mutter näher verwandt
als beim Menschen.
- P(x|a) = 1,
d.h. aus der Sicht des Vaters sind die Töchter näher verwandt
als beim Menschen.
- P(a|x) = 1/2,
d.h. aus der Sicht der Töchter ist der Vater ebenso nahe verwandt
wie beim Menschen.
Die Unsymmetrie
in diesen Beziehungen mag seltsam anmuten, ebenso wie die Formulierung
"aus der Sicht von". Betrachten wir die Sache aber nun "aus
der Sicht eines seltenen Allels", das seine Träger zur Kooperation
auf Kosten eigener Nachkommenschaft veranlasst. Welche Formeln des Verwandtschaftsaltruismus
haben sich eine gute Chance, sich durchsetzen?
Die Grundidee ist dieselbe wie bereits oben diskutiert: Ein Allel, das
c
zum Altruismus gegenüber d
veranlasst, hat umso bessere Chancen auf Ausbreitung, je größer
P(d|c)
ist. Gehen wir die obige Liste durch, so fällt auf, dass die
Schwestern untereinander und mit der Brut, die ja ebenfalls aus
Schwestern besteht, näher verwandt sind als mit der Königin
und mit den eigenen (hypothetischen) Nachkommen! Trägt
eine der Schwestern ein Allel, das zur Unterstützung der Königin
bei der Erzeugung von Nachkommen (d.h. von engen Verwandten) bei gleichzeitigem
Verzicht auf eigenen Nachwuchs veranlasst, so hat dieses gute Aussichten
auf Verbreitung in der gesamten Population. Beachten Sie vor allem die
drei farblich hervorgehobenen Zahlenwerte
der obigen Liste: Da r(x, y) = 3/4,
aber nur r(x, b) = 1/2
und P(z|b) = 1/2,
ist die Wahrscheinlichkeit, dass eine andere Schwester das Allel ebenfalls
trägt, größer als die Wahrscheinlichkeit, es
einem Nachkommen (gleich welchen Geschlechts) zu vererben! Diese Tatsache
erklärt, dass bei Hautflüglern die sterilen Arbeiterkasten
aus Weibchen bestehen. (Staaten mit sterilen Kasten haben sich bei den
Hautflüglern elf mal unabhängig voneinander entwickelt, im
gesamten übrigen Tierreich, so weit wir wissen, nur zweimal, nämlich
bei den Termiten und Nacktmullen, bei denen die Arbeiterkasten aus Individuen
beiderlei Geschlechts bestehen, und auf die wir im übernächsten
Abschnitt zu sprechen kommen).
Wird
der Sache quantitativ näher auf den Grund gegangen, so stellt
sich heraus, dass sie nur dann so richtig in Gang kommt, wenn es den
Schwestern gelingt, die Anzahl der von der Königin produzierten
Söhne (die aus der Sicht der Schwestern nur weitläufige
Verwandte sind) zu reduzieren. Der Grund dafür liegt darin, dass
die Anreicherung des Genpools mit dem Kooperations-Allel durch die
geringe Wahrscheinlichkeit, dass es ein Bruder auch tragen wird, behindert
wird. Aber auch diese Hürde haben die Hautflügler-Schwestern
geschafft.
Eine
weitere theoretische Überlegung, auf die wir hier nicht eingehen
können, zeigt übrigens einen atemberaubenden Interessenskonflikt
auf: Aus der Sicht eines Kooperations-Allels, wenn es sich im Organismus
einer der Schwestern befindet, sollte die Königin nur wenige
Söhne bekommen. Das aus dieser Sicht optimale Töchter-Söhne-Verhältnis
wurde mit 3:1
ermittelt. Ist dasselbe Allel jedoch im Körper einer Königin,
so hat es eher ein Interesse am Verhältnis
1:1! Tatsächlich gibt es Hinweise, dass die 3:1-Variante
bei vielen Arten realisiert ist. In (natürlich unzutreffenden)
menschlichen Begriffen ausgedrückt bedeutet das, dass nicht die
Arbeiterinnen die Sklavinnen der Königin sind, sondern umgekehrt
die Königin als Maschine zur Schwesternproduktion betreiben!
Die in der Literatur beschriebenen Verwicklungen dieser Art können
es durchaus mit einem Kriminalroman aufnehmen...
Auch wenn die Ursprünge
des extrem hohen Kooperationsniveaus der Bienen, Wespen und Ameisen
noch umstritten sind und immer neue Gesichtpunkte zu Tage treten, besteht
Einigkeit, dass es eng mit der - uns Menschen
eigenartig anmutenden - Fortpflanzungsart
dieser Tiere zusammenhängt.
Ressourcen:
Literatur zu Verwandtschaft
und Verwandtschaftsaltruismus:
- Richard Dawkins, Das egoistische
Gen, Rowohlt, 1996
- Karl Sigmund,
Spielpläne, Knaur, 1997
|