 Homepage
Homepage
Present position: Senior Scientist/External Docent/Lecturer at the University of Vienna
Email: katrin.teubner@univie.ac.at
Department: Department of Functional and Evolutionary Ecology
Personal homepage: https://www.lakeriver.at/
Education and degrees
Habilitation in limnology (thesis on ecological stoichiometry), Austria 06/2004, Univ. Vienna, Fac. Sci. an Math., Inst. Ecology and Conserv. Biol.; Ph. D. in ecology (thesis on phytoplankton), GDR, 12/1996, Humboldt Univ. Berlin, Dept. Biology, Inst. Ecophysiology., M. Sc. in education on Biology and Chemistry (thesis on reed belts), GDR, 07/1985 teacher training college, Köthen (Sachsen-Anhalt).
Research interest: My main interest is to advance our understanding on the ecological and physiological response patterns of microbial communities to environmental variability. The main emphasis of my present and future research is on primary producers in aquatic ecosystems.
- Photosynthesis, pigment pattern and nutrient uptake of phytoplankton
- Ecology of aquatic microbial community
- Applied issues: lake restoration, Water Frame Work Directive, Response to climate change
A key reference point for ecological stoichiometry in freshwaters is TN:TP:SRSi = 16:1:17
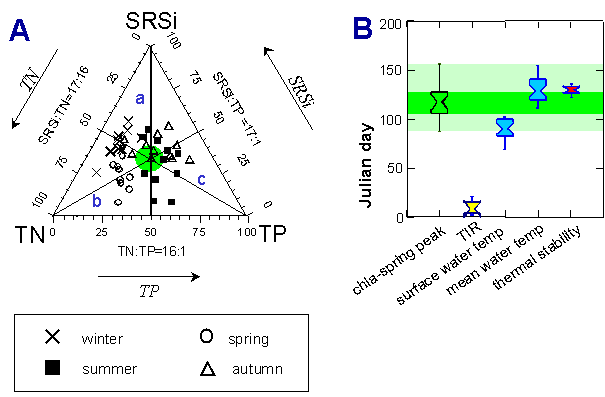
Our concept of the ‘balance of TN: TP: SRSi-ratios’ attempts to determine whether the seasonal succession of phytoplankton communities is influenced by or responds to nutrient stoichiometry (TN = total nitrogen, TP = total phosphorus, SRSi = soluble reactive silica; Teubner & Dokulil 2002). Because the molar average of TN: TP: SRSi = 20: 1: 17 for lakes across shallow to deep, oligotrophic to hypertrophic, clear-water to dystrophic and flushed-riverine to stratified water bodies was close to the marine optimum stoichiometric ratio, we used the ratio N : P : Si = 16 : 1: 17 as key reference point for ecological stoichiometry in freshwaters. Triangular diagrams of TN: TP: SRSi ratios scaled in these physiological proportions of 16: 1: 17 were aimed at synoptically presenting of relative nutrient availability for both diatoms and non-siliceous algae in phytoplankton communities. In lakes with ‘balanced nutrient ratios’ TN: TP: SRSi ratios were evenly fluctuating around the stoichiometric optimum of 16:1:17, in a cyclic pattern within a given year (Fig. 1A, pdf). These lakes with balanced nutrient ratios yielded maximum algal biomass but were more the exception than the rule among the 11 studied water bodies. Our findings strongly support that lakes with one invariant nutrient element in relation to those of the other two display commonly the seasonal pattern of nutrient availability. The unbalanced dynamic of seasonal nutrient ratios were associated with relatively constant concentrations of either SRSi, or TN or TP in a certain lake type. The relative change in seasonal availability of silica, of the element of long turnover times as known from literature, was more effected by distinct lake morphometry than that of TN or TP.
Resource accumulation efficiency
The Redfield ocean can be seen as the ‘perfect sea’ due to balanced flow of C, N and P in and out of the biota. · In the context of stoichiometric ecology, we call a nutrient rich, shallow riverine lake in North-Germany the ‘perfect lake’ because (i) the ratios of ‘external” nutrients, TN:TP:SRSi, behave like in a lake with ‘balanced nutrient ratios’ defined above and (ii) the elemental ratio of biota is very close to C:N:P= 106:16:1 (Teubner, 2004). An overlay of both seasonal patterns, TN:TP:SRSi and POC:PON:POP (= C:N:P of biota), mirrors the complementary relationship between external and biota-internal stoichiometry in this lake (pdf). External P-limitation, indicated by relatively low TP, is accompanied by a P-rich phytoplankton community (dominated by diatoms), in turn, N-limitation by an N-rich assemblage (dominated by cyanobacteria). The stoichiometric shift towards the limiting element is a common phenomenon of individual adaptation/acclimation of producer organisms and can be recognised even at the ecosystem level (Teubner et al. 1999 pdf, Teubner et al. 2003 a, b; Teubner 2003).
Photoacclimation
We found evidence that low-light conditions due to turbidity caused by industrial effluents in deep alpine lake Traunsee were compensated by photoacclimation and therefore did not result in significant quantitative changes of photosynthetic rates and algal biomass (Teubner & Dokulil 2002 b). Our study on photosynthetic properties in three alpine lakes, Lake Lucerne, Mondsee and Traunsee in fall 1999, could further show, that light acclimation were mirrored concurrently by photosynthetic efficiency and pigment pattern and were primarily due to differences in cell size than a different taxonomic composition among natural phytoplankton assemblages (Teubner et al. 2001 link to pdf). Low-light acclimation in natural plankton was associated with high maximum light utilisation coefficient (α*, 14C method) and enhanced chlorophyll-a to photo-protective β-carotene ratios. These effects of low-light acclimation could be attributed to both the phytoplankton assemblages of particular lakes studied under cloudy weather conditions and to the cell-size fraction smaller than 10 μm in all three lakes in general. Algae grown under higher solar radiation of sunny days and of the size fraction larger than 10 μm were photo-synthetically less efficient but high-light acclimated having low ratios of chlorophyll-a to ß-carotene. Above photosynthetic saturating light levels, only the high light acclimated large-cell fraction increased their relative proportion of photo-protective carotenoids. At dim-light in deeper water layers, however, allometric rules seemed to be counterbalanced since the proportion between lipophilic accessory photo-synthetic pigments and photo-protective pigments increased concurrently for both size fractions.
The pigment pattern in epilimnetic strata and the metalimnetic layers of the deep chlorophyll maximum mirrored distinct phytoplankton assemblages due to vertical niche separation in deep mesotrophic lakes (Ammersee, Bavaria: Teubner et al. 2003 b; Mondsee, Salzkammergut Austria: Greisberger & Teubner 2007 pdf). A detailed comparative study on in vivo measurements of delayed fluorescence (DF) and HPLC/CHEMTAX pigment analysis we used to infer seasonal and depth distributions of phytoplankton in relation to light and temperature in deep alpine mesotrophic lake Mondsee (Austria). The advantage of HPLC/CHEMTAX method became evident in the good discrimination between phytoplankton groups based on a pigment ratio matrix derived from multiple regression analysis of phytoplankton samples. Changes in the pattern of photosynthetic active pigments were investigated with DF and benefited from excitation spectra that considered all light harvesting pigments. The enhancement of accessory photosynthetic active pigments relative to active chlorophyll a under ambient low light and temperature became evident along both the seasonal succession and the depth gradients by shifts toward diatoms and/or phycobilin-rich cyanobacteria and cryptophyceen in plankton communities (Greisberger & Teubner, 2007 pdf).
Community ecology
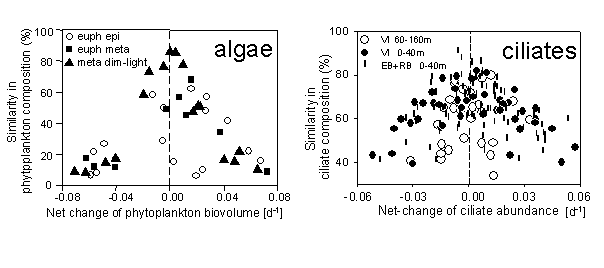
We have shown that both algal (Teubner et al. 2003 b) and ciliate plankton assemblages (Sonntag et al., 2006 pdf) reached the highest persistence in species composition (high similarity between successive samples) during periods of almost constant standing crop (low values of net change of total biovolume) and vice versa (Fig. 2). In this view we define a steady state assemblage as a stable community concerning both the species composition and the standing crop as exemplified by phytoplankton of metalimnetic strata at dim-light levels below 1% in deep alpine lakes, but not for assemblages in euphotic surface layers. Shifts towards a P-rich biota under reduced total-P pool-size in eutrophied Old Danube strongly indicated that the plankton community acted as a sink for phosphorus (Teubner et al. 2003 a pdf). Our hypothesis that the increased P-accumulation in organisms reflects the adaptive P-utilisation of a planktonic community under low P-supply was substantiated by changes in the structure and dynamics on the nutrient-producer and producer-consumer interfaces. These findings of enhanced P-accumulation by plankton community under oligotrophic conditions were verified again for the epilimnion of open waters of the deep alpine lake Traunsee (Teubner 2003).
Decadal trends and global change
Our analysis on six alpine lakes in the Austrian ‘Salzkammergut’ district confirmed the hypothesis that spatial coherence is best demonstrated via physical parameters and less pronounced in the sequence of basic chemical, nutrient and biological variables (Dokulil & Teubner, 2002 a; Dokulil & Teubner, 2005 a, b, Dokulil et al. 2006 pdf). The significant deep water warming for European lakes could shown in Dokulil et al. 2006, including Ammersee and Mondsee (Fig. 2 f and k, pdf).
Applied freshwater science
Through our interest in diatoms (Teubner, 1995 pdf) and cyanobacteria (Teubner, 2006 pdf; Teubner et al., 1999 pdf, 2004 pdf) in particular and ecosystems in general we became involved in applied issues of eutrophication (Dokulil & Teubner, 2000 a, b), and restoration of lakes (Teubner et al., 2003 a, Dokulil et al. 2000), climatic change (Teubner et al., 2006; Dokulil & Teubner 2002 a, Dokulil et al. 2006 pdf) and the Water Framework Directive.
Selected publications (updated Sept. 2022)
- Teubner K, Teubner IE, Pall K, Tolotti M, Kabas W, Drexler S-S, Waidbacher H, Dokulil MT (2022) Macrophyte habitat architecture and benthic-pelagic coupling: Photic habitat demand to build up large P storage capacity and bio-surface by underwater vegetation. Frontiers in Environmental Science, 10:901924. DOI:10.3389/fenvs.2020.573724
- pdf Teubner K, Teubner IE, Pall K, Kabas W, Tolotti M, Ofenböck T, Dokulil MT (2021) New Emphasis on Water Clarity as Socio-Ecological Indicator for Urban Water – a short illustration. In: Rivers and Floodplains in the Anthropocene – Upcoming Challenges in the Danube River Basin, Extended Abstracts 43rdIAD-conference (DOI:10.17904/ku.edoc.28094):70-78
- Dokulil, M. & K. Teubner 2010. Eutrophication and Climate Change: Present Situation and Future Scenarios. In: Eutrophication: Causes, Consequences and Control (eds. A.A Ansari, S.S. Gill, G.R. Lanza, W. Rast) Springer Publisher. in press.
- Dokulil, M., K. Teubner, A. Jagsch, U. Nickus, R. Adrian, D. Straile, T. Jankowski, A. Herzig, A. & J. Padisák 2010. The impact of climate change on lakes in Central Europe. In: The Impact of Climate Change on European Lakes (Ed. DG George), Aquatic Ecology Series (Ed. J. Huisman), Springer Publisher, 387-410.
- Nõges, P., O. Anneville, L. Arvola, T. Blenckner, D.G. George, D.G., T. Jankowski, M. Järvinen, S.C. Maberly, J. Padisák, D. Straile, K. Teubner & G. Weyhenmeyer. 2010. The impact of variations in the climate on seasonal dynamics of phytoplankton. In: The Impact of Climate Change on European Lakes (Ed. DG George), Aquatic Ecology Series (Ed. J. Huisman), Springer Publisher, 253-276.
- Strausz, V., Janauer, G. A. & Teubner, K. 2010. Spatial patchiness and similarity of macrophyte assemblages along a cut-off channel of the River Danube in Linz (Austria). Proceedings IAD conference, Dresden.
- pdf Greisberger, S., Dokulil, M. & K. Teubner. 2008. A comparison of phytoplankton size-fractions in Mondsee, an alpine lake in Austria: distribution, pigment composition and primary production rates. Aquatic Ecology, 42: 379-389.
- pdf Greisberger, S. & K. Teubner. 2007. Does pigment composition reflect phytoplankton community structure in differing temperature and light conditions in a deep alpine lake? An approach using HPLC and delayed fluorescence (DF) techniques. J Phycol, 43, 1108-1119.
- Blenckner, T., Adrian, R., Livingstone, D.M., Jennings, E., G. A. Weyhenmeyer, Aonghusa, C. N., George, D. G., Jankowski, T., Järvinen, M., Nõges, T., Straile, D. &. K. Teubner. 2007. Large-scale climatic signatures in lakes across Europe: a meta-analysis. Global Change Biology, 13: 1314-1326.
- pdf Kaiblinger, C., Greisberger, S., Teubner, K. & M.T. Dokulil. 2007. Photosynthetic efficiency as a function of thermal stratification and phytoplankton size structure in an oligotrophic alpine lake. Hydrobiologia 578: 29-36
- pdf Dokulil, M. T., Jagsch, A., George, G. D., Anneville, O., Jankowski, T., Wahl, B., Lenhart, B., Blenckner T. & K. Teubner. 2006. Twenty years of spatially coherent deep-water warming in lakes across Europe related to North-Atlantic Oscillation. Limnol Oceanogr, 51 (6): 2787-2793.
- pdf Sonntag, B., Posch, T., Klammer, S., Teubner, K. & R. Psenner. 2006. Ciliates and flagellates in an oligotrophic deep alpine lake: contrasting variability with seasons and depths. Aquat Microb Ecol., 43: 193-207.
- pdf Teubner, K. 2006. Ergebnisse des Forschungsvorhabens „Bedingungen für das Auftreten toxinbildender Cyanobakterien (Blaualgen) in bayerischen Seen und anderen stehenden Gewässern. p.49-74. In: Toxinbildende Cyanobakterien (Blaualgen) in bayerischen Gewässern: Massenentwicklungen, Gefährdungspotential, wasserwirtschaftlicher Bezug. ed Ha Morscheid, Bayerisches Landesamt für Umwelt, Materialienband Nr. 125, ISBN: 13: 978-3-940009-08-1
- Dokulil, M.T. & K. Teubner. 2005. The global warming versus re-oligotrophication controversy in lakes: Can effects on phytoplankton be disentangled? Phycologia 44 (Suppl.): 28-29.
- pdf Dokulil, M.T. & K. Teubner. 2005. Do phytoplankton assemblages correctly track trophic changes? – An assessment using directly measured and palaeolimnological data. Freshwater Biol, 50 (10), 1594–1604.
- Teubner, K. 2005. Pigmente. In Primärproduktion (autotrophe Produktion), p 6-9. Handbuch Angewandte Limnologie, Limnologische Grundlagen – Gewässerbelastungen – Restaurierung – Aquatische Ökotoxikologie – Gewässerschutz – Bewertung., eds Steinberg et al. ISBN 3-609-75820.
- pdf Donabaum, K., Pall,K., Teubner, K. & M. t. Dokulil. 2004. Alternative stable states, resilience and hysteresis during recovery from eutrophication – A case study. SIL-News (43), 1-4.
- pdf Teubner, K., Morscheid, Ha., Tolotti, M., Morscheid, Hei. & V. Kucklentz. 2004. Bedingungen für das Auftreten toxinbildender Blaualgen in bayerischen Seen und anderen stehenden Gewässern. Informationsberichte Bayer. Landesamt für Wasserwirtschaft, München: 84 pp.
- pdf Teubner, K., N. Crosbie, K. Donabaum, W. Kabas, A. Kirschner, G. Pfister, M. Salbrechter & M. T. Dokulil. 2003 (a). Enhanced phosphorus accumulation efficiency by the pelagic community at reduced phosphorus supply: a lake experiment from bacteria to metazoan zooplankton. Limnol Oceanogr 48 (3): 1141-1149.
- pdf Teubner, K., Tolotti, M., Greisberger, S., Morscheid, H., Dokulil, M.T. & Morscheid, H. 2003 (b). Steady state phytoplankton in a deep pre-alpine lake: Species and pigments of epilimnetic versus metalimetic assemblages. Hydrobiologia 502: 49-64.
- Teubner, K. 2003. Phytoplankton, pelagic community and nutrients in a deep oligotrophic alpine lake: Ratios as sensitive indicators of the use of P-resources (DRP:DOP:PP and TN:TP:SRSi). Water Res 37 (7): 1583-192.
- pdf Crosbie, N.D., Teubner, K. & T. Weisse. 2003. Flow-cytometric mapping provides novel insights into the seasonal and vertical distributions of freshwater autotrophic picoplankton. Aquat Microb Ecol 33: 53-66.
- Dokulil, M.T. & K. Teubner. 2003. Steady state phytoplankton assemblages during thermal stratification in deep alpine lakes. Do they occur? Hydrobiologia 502: 65-72.
- Chen, Y., Qin Boqiang, Teubner, K. & M. T. Dokulil. 2003. Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China. J Plankton Res 25 (1), 445-453.
- Chen, Y, Chengxin Fan, Teubner, K. & M. Dokulil. 2003. Changes of nutrients and phytoplankton chlorophyll-a in a large shallow lake, Taihu, China: an 8-year investigation. Hydrobiologia 506: 273-279.
- Dokulil, M. & K. Teubner. 2003. Eutrophication and restoration of shallow lakes – the concept of stable equilibria revisited. Hydrobiologia 506: 29-35.
- Dokulil, M. & K. Teubner. 2003. Klimaeinflüsse auf Seen in Europa (CLIME). Österr. Fischerei 56: 176-180.
- Teubner, K., & M. T. Dokulil. 2002. 14C-Photosynthesis of phytoplankton in an oligotrophic alpine lake (Traunsee, Austria) and its response to turbidity caused by industrial tailings. Water Air Soil Poll, Focus 2: 181-190.
- pdf Teubner, K. & M. T. Dokulil. 2002. Ecological stoichiometry of TN:TP:SRSi in freshwaters: nutrient ratios and seasonal shifts in phytoplankton assemblages. Arch Hydrobiol. 154 (84): 625-646.
- link to pdf Teubner, K., A. Sarobe Egiguren, M. R. Vadrucci & M. Dokulil. 2001. 14C photosynthesis and pigment pattern of phytoplankton as size related adaptation strategies in alpine lakes. Aquat Sci 63: 310-325.
- Teubner, K. 2001. Algengemeinschaften in Seen. 83-112. In: Ökologie und Schutz von Seen. UTB Facultas, Wien.
- pdf Teubner, K. 2000. Synchronised changes of planktonic cyanobacterial and diatom assemblages in North German waters reduce seasonality to two principal periods. Arch Hydrobiol, Spec Issues Advanc Limnol 55: 564-580.
- Dokulil, M. & K. Teubner. 2000. Cyanobacterial dominance in lakes. Hydrobiologia 438: 1-12.
- Dokulil, M. & K. Teubner. 2000. Effects of reduced load in the river Danube on nutrients and phytoplankton dynamics in the flood-relief channel New Danube, Vienna Austria. Arch Hydrobiol, Suppl. Large Rivers 12 (1): 39-59.
- Dokulil, M. T., K. Teubner & K. Donabaum. 2000. Restoration of a shallow, groundwater fed urban lake using a combination of internal management strategies: a case study. Arch Hydrobiol, Spec Issues Advanc Limnol 55: 271-282.
- pdf Teubner, K., R. Feyerabend, M. Henning, A. Nicklisch, P. Woitke & J.-G. Kohl. 1999. Alternative blooming of Aphanizomenon flos-aquae or Planktothrix agardhii induced by the timing of the critical nitrogen : phosphorus ratio in hypertrophic riverine lakes. Arch Hydrobiol, Spec Issues Advanc Limnol 54: 325-344.
- pdf Teubner, K. 1997. Merkmalsvariabilität bei planktischen Diatomeen in Berlin-Brandenburger Gewässern. Nova Hedwigia 65 (1-4): 233-250.
- pdf Teubner, K. 1995. A light microscopical investigation and multivariate statistical analyses of heterovalvar cells of Cyclotella-species (Bacillariophyceae) from lakes of the Berlin-Brandenburg region. Diatom Research 10 (1): 191-205.
Theses
- Teubner, K. 2004. More or less? Smaller or bigger? How relevant are relative changes in aquatic ecosystems? Habilitation thesis on Ecological Stoichiometry, Fac. of Sciences and Mathematics, Institute of Ecology and Conservation Biology University Vienna, 188 pp.
- pdf Teubner, K. 1996. Struktur und Dynamik des Phytoplanktons in Beziehung zur Hydrochemie und Hydrophysik der Gewässer: Eine multivariate statistische Analyse an ausgewählten Gewässern der Region Berlin-Brandenburg. Ph.D thesis, Dept. Ecophysiology, Humboldt University Berlin, 232 pp.
- Pabst, K. 1985. Ökologische Untersuchungen als Grundlage für landeskulturelle Maßnahmen im Bereich der Michelner Teiche (Kreis Köthen) – Die Pflanzengesellschaften der Wasser-, Ufer- und Verlandungsbereiche. M.Sc. thesis, Dept. Biology and Chemistry, Teacher training college (Pädagogische Hochschule ‘W. Ratke’), Köthen/Sachsen-Anhalt, Germany.